Биологическая роль мембранных белков. Клеточная мембрана: определение, функции мембран. Жидкостно-кристаллическая модель. Количественные пропорции мембранных белков и липидов. Функции мембранных белков, липидов, углеводов. Латеральная диффузия белков, лип
В настоящее время очень активно обсуждается персонифицированная медицина и методы точечного и селективного воздействия на отдельные молекулы и системы клеток, ответственные за развитие тех или иных заболеваний. Мембранные белки при этом выделяются как основной класс возможных мишеней для терапии, так как именно они обеспечивают непосредственную передачу сигналов в живую клетку. Можно отметить, что в настоящее время более половины всех лекарственных препаратов имеют мишень на клеточной мембране, а дальше таких соединений будет становиться только больше.
Структура плазматической мембраны
Думаю, многие помнят из школьного курса биологии, как устроена живая клетка. И несомненно, особое место в устройстве клетки занимает плазматическая мембрана. Именно она отделяет внутриклеточное пространство от внеклеточного и является границей клетки. В соответствии с таким представлением основной функцией биологической мембраны является создание барьера между «живым» внутриклеточным пространством и окружающей средой. Последнюю можно условно считать «неживой» для одноклеточных организмов, например бактерий. Клетки бактерий, архей, грибов и растений покрыты поверх мембраны еще и более прочной жесткой оболочкой - клеточной стенкой. Она также служит для защиты клетки от внешних воздействий. Однако у плазматической мембраны существует еще множество других функций.
В 1925 году И. Гортер и А. Грендель показали, что клеточная мембрана представляет собой двойной слой (бислой) из молекул липидов. Основной их чертой является амфифильность, то есть наличие в молекуле двух частей с разными свойствами. Так, выделяют гидрофильную (буквально ‘любящую воду’) полярную «головку» и липофильные (буквально ‘любящие жиры’) ацильные цепи. При образовании мембраны липофильные участки молекул оказываются обращенными внутрь бислоя, а гидрофильные - наружу. Такая схема организации мембраны характерна для большинства организмов, поэтому существует множество биомолекул, «нацеленных» на непосредственное взаимодействие с липидным бислоем. Следует отметить, что мембраны органелл клеток образованы по такому же принципу, однако состав липидов в них отличается от плазматической мембраны.
В 1935 году Дж. Даниэлли и Х. Доусон показали, что в клеточной мембране, помимо липидов, содержатся белки. Так возникла модель «сэндвича», в которой плазматическая мембрана представлялась в виде двух слоев белков, между которыми располагался липидный бислой. Эта модель подтверждалась первыми экспериментами по электронной микроскопии мембран, поэтому в 1960 году Дж. Робертсон постулировал одинаковое трехслойное строение для всех мембран живой клетки.
Однако данная модель не могла объяснить накапливающиеся экспериментальные данные, и в 1972 году С. Д. Сингером и Г. Л. Николсоном была предложена жидкостно-мозаичная модель мембраны, где мембранные белки «плавают» в жидком липидном бислое, как айсберги в открытом море. При этом предполагалось, что белки никак не упорядочены и могут свободно перемещаться в мембране. Белки, согласно этой модели, могут как взаимодействовать с поверхностью мембраны и потому находиться с одной ее стороны (периферические белки), так и пронизывать мембрану насквозь (интегральные мембранные белки). Последние имеют, таким образом, возможность взаимодействовать как со внеклеточной средой, так и с цитоплазмой клетки. Иногда также выделяют полуинтегральные белки, частично погруженные в мембрану, но не пронизывающие ее насквозь.
Тем не менее даже такая сложная модель организации биологической мембраны потребовала уточнения, когда к 80-м годам XX века развилась концепция липидных рафтов. Сам термин «липидные рафты» был впервые предложен в 1988 году К. Симонсом и Г. ван Меером для описания выделенных участков плотно упакованного липида. В настоящее время существование липидных доменов (то есть областей с определенным набором свойств) в плазматической мембране подтверждено огромным количеством исследований. Показано, что их формирование определяется в том числе белками, находящимися в рассматриваемой части мембраны. Мембранные белки, таким образом, распределены по поверхности клетки не хаотично, а занимают определенные участки, в которых может достигаться высокоупорядоченная структура.
Мембранные белки
Надо понимать, что живая клетка лишена привычных для нас органов чувств, которые, между прочим, сами состоят из определенных типов клеток. Тем не менее, как и нам с вами, клетке нужно взаимодействовать с окружающей средой. При этом для некоторых воздействий, например для света или для маленьких липофильных молекул, плазматическая мембрана не является препятствием, и потому они могут напрямую взаимодействовать с внутриклеточными белками. Здесь следует напомнить, что в ответ на внешние воздействия в клетке происходят каскады химических реакций, завершающиеся, например, началом выработки определенных белков или запуском определенных программ жизнедеятельности клетки. Так, в ответ на некоторые воздействия клетка может выделять во внешнюю среду гормоны или ферменты, начинать деление или даже запускать запрограммированный механизм собственной гибели - апоптоз. Это далеко не все возможные ответы, однако все они имеют общий принцип запуска в виде каскада химических превращений во внутриклеточном пространстве.
Также для поддержания жизнедеятельности необходим постоянный транспорт вещества через мембрану. Так как существует множество различных внешних сигналов, на которые клетка должна уметь отвечать, на ее поверхности представлено большое многообразие различных мембранных белков. Среди них выделяют рецепторы, ионные каналы, порины, транспортеры, молекулярные моторы и структурные белки. Рецепторные белки формируют внутри клетки сигнал в ответ на появление снаружи гормонов и сигнальных молекул. К ним относится, например, рецептор инсулина, отвечающий за поступление в клетку глюкозы. Ионные каналы обеспечивают транспорт ионов и поддержание градиента (то есть разницы в концентрациях) их концентраций между внешней средой и цитоплазмой клетки. Натриевые и калиевые каналы непосредственно участвуют в передаче нервного импульса. Порины и транспортеры обеспечивают перенос воды и определенных молекул через мембрану. Молекулярные моторы присутствуют у многих бактерий и обеспечивают подвижность клеток. Наконец, структурные белки поддерживают структуру мембраны и взаимодействуют с остальными белками. Не менее сложной является сеть внутриклеточных путей передачи сигналов с помощью каскадов реакций. Взаимодействиями белков в клетке и, соответственно, путями передачи сигналов занимается специальное научное направление, называемое интерактомикой (от англ. interaction - ‘взаимодействие’).
Мембрана как среда для взаимодействия белков
Выше уже было рассказано о различных классах мембранных белков и их распределении в плазматической мембране. Существующие представления о строении мембраны говорят о том, что на поверхности клетки формируются отдельные домены, содержащие определенный набор белков и липидов и предназначенные для выполнения заданных функций. Такое пространственное разделение может быть необходимо ввиду сложной организации процессов межмолекулярных взаимодействий и регуляции жизни живой клетки. Естественно, белки не работают поодиночке. Многие из них просто неспособны выполнять свои функции без своих партнеров. Нарушения в этом сложном механизме могут приводить к тому, что клетка начнет неправильно реагировать на внешние сигналы. Это приводит к развитию таких серьезных заболеваний, как рак и диабет, а также к целому ряду других. Для их лечения мы должны понимать, как работают эти сигнальные системы клетки и как можно на них воздействовать. Природа сама предлагает нам множество инструментов для такого воздействия: молекулы гормонов, а также токсины из различных ядов способны прицельно выбирать свои мишени на поверхности плазматической мембраны.
Однако далеко не всегда их можно использовать в исходном виде, поэтому одной из задач современной молекулярной биологии является разработка новых модифицированных соединений, которые бы селективно действовали на мембранные мишени и в то же время были лишены недостатков исходных природных молекул. При таких исследованиях необходимо понимание не только механизма узнавания между мишенью и терапевтическим агентом, но и всей специфики взаимной организации мембранных белков и мембраны в целом.
Надо понимать, что мембрана - это не просто инертная матрица, которая нужна для того, чтобы белки в ней расположились и работали. Мембрана - это очень активная и динамичная среда, которая существенным образом влияет на работу находящихся в ней белков. Это открывает новые пути для воздействия на мембранные белки путем изменения свойств самого липидного бислоя. Также крайне важным является взаимодействие между трансмембранными доменами белков. Оно определяет правильную организацию и функционирование огромного ряда жизненно важных систем клетки. Таким образом, мы должны иметь представление не только о функциях конкретных белков, но и о том, какие существуют взаимосвязи между их работой и свойствами мембраны. Первые результаты в этом направлении уже получены, однако на данном этапе мы еще не имеем перед собой полной картины функционирования мембраны клетки.
Рецепторные системы клетки
Среди всех мембранных белков, о которых было сказано выше, особое положение занимают рецепторы. Они обеспечивают передачу информации через плазматическую мембрану внутрь клетки. У многоклеточных организмов (и у многих бактерий, образующих колонии) передача информации между клетками осуществляется с помощью специальных сигнальных молекул, выделяемых во внешнюю среду. Среди этих сигнальных молекул выделяют гормоны - молекулы белковой природы, которые специально секретируются (выделяются) в специализированных органах. Также в качестве сигнальных молекул могут выступать небелковые соединения и даже ионы, образующиеся или высвобождающиеся, например, при повреждении соседних клеток. Таким образом устроена передача болевых сигналов, выполняющая важнейшую защитную функцию в организме. Рецепторы специфически распознают сигналы (молекулы лигандов), появляющиеся во внеклеточном пространстве, и запускают каскады реакций в ответ на них. Можно выделить два ключевых класса рецепторов: рецепторы, сопряженные с G-белком (GPCR), и рецепторные тирозинкиназы (РТК).
Рецепторные тирозинкиназы - это обширный класс мембранных белков. К ним относятся рецепторы целого ряда факторов роста и гормонов. Одним из наиболее известных белков этого класса является рецептор инсулина. Без него был бы невозможен управляемый транспорт глюкозы в клетке, а нарушения в работе этого белка приводят к развитию диабета II типа. Рецепторы эпидермального фактора роста оказываются еще более важными, поскольку регулируют клеточный цикл. Неправильная их работа зачастую становится причиной развития онкологических заболеваний. Большинство из этих рецепторов имеют, на первый взгляд, очень простую структуру: один внеклеточный домен, одну трансмембранную альфа-спираль и один цитоплазматический киназный домен. Однако не все так просто, и поодиночке РТК не работают. Для активации им необходимо сформировать пару (или, по-научному, димер). Гормон или фактор роста взаимодействует со внеклеточным доменом РТК и способствует его димеризации, то есть образованию димера со вторым таким же рецептором. После этого происходит димеризация в трансмембранном и киназном доменах. В результате димер киназного домена становится отправной точкой для каскада химических реакций. Здесь описано лишь общее представление о механизме работы РТК, поскольку детали этого процесса довольно сложны и не до конца понятны даже по сей день.
К GPCR относятся зрительный родопсин, вкусовые рецепторы, рецепторы запахов и многие другие. Это большие молекулы, интегрированные в плазматическую мембрану. Они содержат по семь трансмембранных альфа-спиралей, которые пронизывают мембрану как столбы. В ответ на связывание лиганда снаружи мембраны эти белки претерпевают конформационную перестройку таким образом, что начинают взаимодействовать с G-белком, расположенным вблизи внутренней поверхности плазматической мембраны. При этом исходный сигнал может быть усилен, поскольку активированный рецептор может активировать несколько молекул G-белка.
Фармакологические мишени
В предыдущем разделе мы говорили о рецепторных системах клетки. Поскольку через них идет большинство сигналов, то именно они и являются теми точками, на которые мы можем воздействовать с помощью фармакологических средств. Действительно, более половины существующих лекарственных препаратов «нацелены» на мембранные белки. Они могут как блокировать их связывание с соответствующими лигандами, так и, наоборот, вызывать активацию и передачу соответствующего сигнала. Второй традиционной мишенью для лекарственных средств и природных токсинов являются ионные каналы.
Основной проблемой здесь является создание лекарственных соединений, обладающих очень высокой селективностью, то есть избирательно воздействующих только на один тип рецептора или даже на определенный его подтип. Это позволяет, например, оказывать воздействие только на определенные типы клеток в организме, поскольку в разных клетках, в зависимости от специализации, на мембране могут быть представлены несколько разные рецепторы. И селективное действие только на определенный их подтип может, например, помочь отделить опухолевые клетки от здоровых.
Это свойство в настоящее время используется в концепции создания модульных структур, когда фармакологическое средство представляет собой несколько молекул (или даже наночастиц), объединенных («сшитых») между собой. При этом возможно совмещение в одном лекарстве нескольких различных по своей функции частей. Так, можно создать «нацеливающую» часть, узнающую на поверхности клетки определенные рецепторы, связанные, например, с развитием рака. К ней добавляется мембранолитический (то есть разрушающий мембрану) агент либо проникающий через мембрану блокатор процессов транскрипции или трансляции (то есть останавливается процесс производства белков в клетке). В результате получается высокоэффективное фармакологическое средство, проявляющее минимум побочных эффектов. Первые лекарства нового поколения уже проходят стадию клинических испытаний и скоро будут применяться для высокоэффективной терапии.
Классификация
Мембранные белки могут быть классифицированы по топологическому или биохимическому принципу. Топологическая классификация основана на локализации белка по отношению к липидному бислою. Биохимическая классификация основана на прочности взаимодействия белка с мембраной.
Различные категории политопических белков. Связывание с мембраной за счёт (1) единичной трансмембранной альфа-спирали, (2) множественных трансмембранных альфа-спиралей, (3) бета-складчатой структуры.

Различные категории интегральных монотопических белков. Связывание с мембраной за счёт (1) амфипатической альфа-спирали, параллельной плоскости мембраны, (2) гидрофобной петли, (3) ковалентно соединённого жирнокислотного остатка, (4) электростатического взаимодействия (прямого или кальций -опосредованного).
Топологическая классификация
По отношению к мембране мембранные белки делятся на поли- и монотопические.
- Политопические, или трансмембранные, белки полностью пронизывают мембрану и, таким образом, взаимодействуют с обеими сторонами липидного бислоя. Как правило, трансмембранный фрагмент белка является альфа-спиралью , состоящей из гидрофобных аминокислот (возможно от 1 до 20 таких фрагментов). Только у бактерий , а также в митохондриях и хлоропластах трансмембранные фрагменты могут быть организованы как бета-складчатая структура (от 8 до 22 поворотов полипептидной цепи).
- Интегральные монотопические белки постоянно встроены в липидный бислой, но соединены с мембраной только на одной стороне, не проникая на противоположную сторону.
Биохимическая классификация
По биохимической классификации мембранные белки делятся на интегральные и периферические .
- Интегральные мембранные белки прочно встроены в мембрану и могут быть извлечены из липидного окружения только с помощью детергентов или неполярных растворителей. По отношению к липидному бислою интегральные белки могут быть трансмембранными политопическими или интегральными монотопическими.
- Периферические мембранные белки являются монотопическими белками. Они либо связаны слабыми связями с липидной мембраной, либо ассоциируют с интегральными белками за счёт гидрофобных, электростатических или других нековалентных сил. Таким образом, в отличие от интегральных белков они диссоциируют от мембраны при обработке соответствующим водным раствором (например, с низким или высоким pH, с высокой концентрацией соли или под действием хаотропного агента). Эта диссоциация не требует разрушения мембраны.
Мембранные белки могут быть встроены в мембрану за счёт жирнокислотных или пренильных остатков либо гликозилфосфатидилинозитола , присоединённых к белку в процессе их посттрансляционной модификации .
Ссылки
Wikimedia Foundation . 2010 .
9.5.1. Одна из главных функций мембран - участие в переносе веществ. Этот процесс обеспечивается при помощи трёх основных механизмов: простой диффузией, облегчённой диффузией и активным транспортом (рисунок 9.10). Запомните важнейшие особенности этих механизмов и примеры транспортируемых веществ в каждом случае.
Рисунок 9.10. Механизмы транспорта молекул через мембрану
Простая диффузия - перенос веществ через мембрану без участия специальных механизмов. Транспорт происходит по градиенту концентрации без затраты энергии. Путём простой диффузии транспортируются малые биомолекулы - Н2 О, СО2 , О2 , мочевина, гидрофобные низкомолекулярные вещества. Скорость простой диффузии пропорциональна градиенту концентрации.
Облегчённая диффузия - перенос веществ через мембрану при помощи белковых каналов или специальных белков-переносчиков. Осуществляется по градиенту концентрации без затраты энергии. Транспортируются моносахариды, аминокислоты, нуклеотиды, глицерол, некоторые ионы. Характерна кинетика насыщения - при определённой (насыщающей) концентрации переносимого вещества в переносе принимают участие все молекулы переносчика и скорость транспорта достигает предельной величины.
Активный транспорт - также требует участия специальных белков-переносчиков, но перенос происходит против градиента концентрации и поэтому требует затраты энергии. При помощи этого механизма через клеточную мембрану транспортируются ионы Na+ , K+ , Ca2+ , Mg2+ , через митохондриальную - протоны. Для активного транспорта веществ характерна кинетика насыщения.
9.5.2. Примером транспортной системы, осуществляющей активный транспорт ионов, является Na+ ,K+ -аденозинтрифосфатаза (Na+ ,K+ -АТФаза или Na+ ,K+ -насос). Этот белок находится в толще плазматической мембраны и способен катализировать реакцию гидролиза АТФ. Энергия, выделяемая при гидролизе 1 молекулы АТФ, используется для переноса 3 ионов Na+ из клетки во внеклеточное пространство и 2 ионов К+ в обратном направлении (рисунок 9.11). В результате действия Na+ ,K+ -АТФазы создаётся разность концентраций между цитозолем клетки и внеклеточной жидкостью. Поскольку перенос ионов неэквивалентен, то возникает разность электрических потенциалов. Таким образом, возникает электрохимический потенциал, который складывается из энергии разности электрических потенциалов Δφ и энергии разности концентраций веществ ΔС по обе стороны мембраны.

Рисунок 9.11. Схема Na+ , K+ -насоса.
9.5.3. Перенос через мембраны частиц и высокомолекулярных соединений
Наряду с транспортом органических веществ и ионов, осуществляемым переносчиками, в клетке существует совершенно особый механизм, предназначенный для поглощения клеткой и выведения из неё высокомолекулярных соединений при помощи изменения формы биомембраны. Такой механизм называют везикулярным транспортом .

Рисунок 9.12. Типы везикулярного транспорта: 1 - эндоцитоз; 2 - экзоцитоз.
При переносе макромолекул происходит последовательное образование и слияние окружённых мембраной пузырьков (везикул). По направлению транспорта и характеру переносимых веществ различают следующие типы везикулярного транспорта:
Эндоцитоз (рисунок 9.12, 1) — перенос веществ в клетку. В зависимости от размера образующихся везикул различают:
а) пиноцитоз — поглощение жидкости и растворённых макромолекул (белков, полисахаридов, нуклеиновых кислот) с помощью небольших пузырьков (150 нм в диаметре);
б) фагоцитоз — поглощение крупных частиц, таких, как микроорганизмы или обломки клеток. В этом случае образуются крупные пузырьки, называемые фагосомами диаметром более 250 нм.
Пиноцитоз характерен для большинства эукариотических клеток, в то время как крупные частицы поглощаются специализированными клетками - лейкоцитами и макрофагами. На первой стадии эндоцитоза вещества или частицы адсорбируются на поверхности мембраны, этот процесс происходит без затраты энергии. На следующей стадии мембрана с адсорбированным веществом углубляется в цитоплазму; образовавшиеся локальные впячивания плазматической мембраны отшнуровываются от поверхности клетки, образуя пузырьки, которые затем мигрируют внутрь клетки. Этот процесс связан системой микрофиламентов и является энергозависимым. Поступившие в клетку пузырьки и фагосомы могут сливаться с лизосомами. Содержащиеся в лизосомах ферменты расщепляют вещества, содержащиеся в пузырьках и фагосомах до низкомолекулярных продуктов (аминокислот, моносахаридов, нуклеотидов), которые транспортируются в цитозоль, где они могут быть использованы клеткой.
Экзоцитоз (рисунок 9.12, 2) — перенос частиц и крупных соединений из клетки. Этот процесс, как и эндоцитоз, протекает с поглощением энергии. Основными разновидностями экзоцитоза являются:
а) секреция - выведение из клетки водорастворимых соединений, которые используются или воздействуют на другие клетки организма. Может осуществляться как неспециализированными клетками, так и клетками эндокринных желёз, слизистой желудочно-кишечного тракта, приспособленными для секреции производимых ими веществ (гормонов, нейромедиаторов, проферментов) в зависимости от определённых потребностей организма.
Секретируемые белки синтезируются на рибосомах, связанных с мембранами шероховатого эндоплазматического ретикулума. Затем эти белки транспортируются к аппарату Гольджи, где они модифицируются, концентрируются, сортируются, и затем упаковываются в пузырьки, которые отщепляются в цитозоль и в дальнейшем сливаются с плазматической мембраной, так что содержимое пузырьков оказывается вне клетки.
В отличие от макромолекул, секретируемые частицы малых размеров, например, протоны, транспортируются из клетки при помощи механизмов облегчённой диффузии и активного транспорта.
б) экскреция - удаление из клетки веществ, которые не могут быть использованы (например, удаление в ходе эритропоэза из ретикулоцитов сетчатой субстанции, представляющей собой агрегированные остатки органелл). Механизм экскреции, по-видимому, состоит в том, что вначале выделяемые частицы оказываются в цитоплазматическом пузырьке, который затем сливается с плазматической мембраной.
Липидам в составе мембран отводят, в первую очередь, структурные свойства - они создают бислой, или матрикс, в котором размещаются активные компоненты мембраны - белки. Именно белки придают разнообразным мембранам уникальность и обеспечивают специфические свойства. Многочисленные мембранные белки выполняют следующие основные функции: обусловливают перенос веществ через мембраны (транспортные функции), осуществляют катализ, обеспечивают процессы фото- и окислительного фосфорилирования, репликацию ДНК, трансляцию и модификацию белков, рецепцию сигналов и передачу нервного импульса и др.
Принято делить мембранные белки на 2 группы: интегральные (внутренние) и периферические (наружные). Критерием такого разделения служит степень прочности связывания белка с мембраной и, соответственно, степень жесткости обработки, необходимой для извлечения белка из мембраны. Так, периферические белки могут высвобождаться в раствор уже при промывке мембран буферными смесями с низкой ионной силой, низкими значениями рН в присутствии хелатирующих веществ, например этилендиаминотетраацетата (ЭДТА), связывающих двухвалентные катионы. Периферические белки выделяются из мембран при таких мягких условиях, поскольку связаны с головками липидов или с другими белками мембраны при помощи слабых электростатических взаимодействий, либо с помощью гидро-фобных взаимодействий - с хвостами липидов. Наоборот, интегральные белки представляют собой амфифильные молекулы, имеют на своей поверхности большие гидрофобные участки и располагаются внутри мембраны, поэтому для их извлечения требуется разрушить бислой. Для этих целей наиболее часто используют детергенты или органические растворители. Способы прикрепления белков к мембране довольно разнообразны (рис. 4.8).
Транспортные белки . Липидный бислой является непроницаемым барьером для большинства водорастворимых молекул и ионов, и их перенос через биомембраны зависит от деятельности транспортных белков. Можно выделить два основных типа этих белков: каналы (поры) и переносчики . Каналы представляют собой туннели, пересекающие мембрану, в которых места связывания транспортируемых веществ доступны на обеих поверхностях мембраны одновременно. Каналы в процессе транспорта веществ не претерпевают каких-либо конформационных изменений, их конформация меняется лишь при открывании и закрывании. Переносчики, наоборот, в процессе переноса веществ через мембрану изменяют свою конформацию. Причем в каждый конкретный момент времени место связывания переносимого вещества в переносчике доступно только на одной поверхности мембраны.
Каналы, в свою очередь, можно разделить на две основные группы: потенциалзависимые и регулируемые химически. Примером потенциалзависимого канала является Na + -канал, его работа регулируется изменением напряжения электрического поля. Иными словами, эти каналы открываются и закрываются в ответ на изменение трансмембранного потенциала . Химически регулируемые каналы
открываются и закрываются в ответ на связывание специфических химических агентов. Например, никотиновый ацетилхолиновый рецептор при связывании с ним нейромедиатора переходит в открытую конформацию и пропускает одновалентные катионы (подрадел 4.7 данной главы). Термины «пора» и «канал» обычно взаимозаменяемы, но под порой чаще понимают неселективные структуры, различающие вещества главным образом по размеру и пропускающие все достаточно малые молекулы. Под каналами чаще понимают ионные каналы. Скорость транспорта через открытый канал достигает 10 6 - 10 8 ионов в секунду.
Переносчики также можно разделить на 2 группы: пассивные и активные. С помощью пассивных переносчиков через мембрану осуществляется транспорт одного типа веществ. Пассивные переносчики задействованы в облегченной диффузии и лишь увеличивают поток вещества, осуществляемый по электрохимическому градиенту (например, перенос глюкозы через мембраны эритроцитов). Активные переносчики транспортируют вещества через мембрану с затратами энергии. Эти транспортные белки накапливают вещества на одной из сторон мембраны, перенося их против электрохимического градиента. Скорость транспорта с помощью переносчиков в очень сильной степени зависит от их типа и колеблется от 30 до 10 5 с -1 . Часто для обозначения отдельных переносчиков используют термины «пермеаза», «транслоказа», которые можно считать синонимами термина «переносчик».
Ферментные функции мембранных белков . В клеточных мембранах функционирует большое количество разнообразных ферментов. Одни из них локализуются в мембране, находя там подходящую среду для превращения гидрофобных соединений, другие благодаря участию мембран располагаются в них в строгой очередности, катализируя последовательные стадии жизненно важных процессов, третьи нуждаются в содействии липидов для стабилизации своей конформации и поддержания активности. В биомембранах обнаружены ферменты - представители всех известных классов. Они могут пронизывать мембрану насквозь, присутствовать в ней в растворенной форме или, являясь периферическими белками, связываться с мембранными поверхностями в ответ на какой-либо сигнал. Можно выделить следующие характерные типы мембранных ферментов:
1) трансмембранные ферменты, катализирующие сопряженные реакции на противоположных сторонах мембраны. Эти ферменты имеют, как правило, несколько активных центров, размещающихся на противоположных сторонах мембраны. Типичными представителями таких ферментов являются компоненты дыхательной цепи или фотосинтетические редокс-центры, катализирующие окислительно-восстановительные процессы, связанные с транспортом электронов и созданием ионных градиентов на мембране;
2) трансмембранные ферменты, участвующие в транспорте веществ. Транспортные белки, сопрягающие перенос вещества с гидролизом АТР, например, обладают каталитической функцией;
3) ферменты, катализирующие превращение связанных с мембраной субстратов. Эти ферменты участвуют в метаболизме мембранных компонентов: фосфолипидов, гликолипидов, стероидов и др.
4) ферменты, участвующие в превращениях водорастворимых субстратов. С помощью мембран, чаще всего в прикрепленном к ним состоянии, ферменты могут концентрироваться в тех областях мембран, где содержание их субстратов наибольшее. Например, ферменты, гидролизующие белки и крахмал, прикрепляются к мембранам микроворсинок кишечника, что способствует увеличению скорости расщепления этих субстратов.
Белки цитоскелета . Цитоскелет представляет собой сложную сеть белковых волокон разного типа и присутствует только в эукариотических клетках. Цитоскелет обеспечивает механическую опору для плазматической мембраны, может определять форму клетки, а также местоположение органелл и их перемещение при митозе. С участием цитоскелета осуществляются также такие важные для клетки процессы, как эндо- и экзоцитоз, фагоцитоз, амебоидное движение. Таким образом, цитоскелет является динамическим каркасом клетки и определяет ее механику.
Цитоскелет формируется из волокон трех типов:
1) микрофиламенты (диаметр ~ 6 нм). Представляют собой нитевидные органеллы - полимеры глобулярного белка актина и других связанных с ним белков;
2) промежуточные филаменты (диаметр 8- 10 нм). Сформированы кератинами и родственными им белками;
3) микротрубочки (диаметр ~ 23 нм) - длинные трубчатые структуры.
Состоят из глобулярного белка тубулина, субъединицы которого формируют полый цилиндр. Длина микротрубочек может достигать нескольких микрометров в цитоплазме клеток и нескольких миллиметров в аксонах нервов.
Перечисленные структуры цитоскелета пронизывают клетку в разных направлениях и тесно связываются с мембраной, прикрепляясь к ней в некоторых точках. Эти участки мембраны играют важную роль в межклеточных контактах, с их помощью клетки могут прикрепляться к субстрату. Они же играют важную роль в трансмембранном распределении липидов и белков в мембранах.
Будущее медицины - персонифицированные методы избирательного воздействия на отдельные системы клетки, которые ответственны за развитие и течение конкретного заболевания. Основным классом терапевтических мишеней при этом являются мембранные белки клетки как структуры, ответственные за обеспечение непосредственной передачи сигналов в клетку. Уже сегодня почти половина лекарств воздействуют именно на клеточные мембраны, и дальше их будет только больше. Знакомству с биологической ролью мембранных белков посвящена данная статья.
Структура и функции клеточной мембраны
Из школьного курса многие помнят устройство структурной единицы организма - клетки. Особое место в устройстве живой клетки играет плазмалемма (мембрана), которая отделяет внутриклеточное пространство от окружающей ее среды. Таким образом, главная ее функция - создание барьера между клеточным содержимым и внеклеточным пространством. Но это не единственная функция плазмолеммы. Среди других функций мембраны, связанных в первую очередь с мембранными белками, выделяют:
- Защитную (связывание антигенов и предупреждение их проникновения в клетку).
- Транспортную (обеспечение обмена веществ между клеткой и средой).
- Сигнальную (встроенные рецепторные белковые комплексы обеспечивают раздражимость клетки и ее ответ на различные воздействия извне).
- Энергетическую - преобразование разных форм энергий: механической (жгутики и реснички), электрической (нервный импульс) и химической (синтез молекул аденозинтрифосфорной кислоты).
- Контактную (обеспечение связи между клетками при помощи десмосом и плазмодесм, а также складок и выростов плазмолеммы).

Строение мембран
Мембрана клетки - это двойной слой липидов. Бислой образуется благодаря наличию в молекуле липидов двух частей с разными свойствами - гидрофильного и гидрофобного участка. Наружный слой мембран образован полярными «головками» с гидрофильными свойствами, а гидрофобные «хвосты» липидов обращены внутрь бислоя. Кроме липидов, в структуру мембран входят белки. В 1972 году американские микробиологи С.Д. Сингер (S. Jonathan Singer) и Г.Л. Николсон (Garth L. Nicolson) предложили жидкостно-мозаичную модель строения мембраны, согласно которой, мембранные белки «плавают» в бислое липидов. Эта модель была дополнена немецким биологом Каем Зимонсом (1997) в части образования определенных, более плотных участков с ассоциированными белками (липидных рафтов), которые свободно дрейфуют в бислое мембраны.
Пространственная структура мембранных белков
В различных клетках соотношение липидов и белков различно (от 25 до 75% белков в пересчете на сухую массу), и расположены они неравномерно. По расположению белки могут быть:
- Интегральными (трансмембранными) - встроенными в мембрану. При этом они пронизывают мембрану, иногда неоднократно. Их внеклеточные участки часто несут цепи олигосахаридов, формируя гликопротеиновые кластеры.
- Периферическими - расположены преимущественно на внутренней стороне мембран. Связь с липидами мембраны обеспечивается за счет водородных обратимых связей.
- Заякоренными - преимущественно расположены с наружной стороны клетки и «якорем», удерживающим их на поверхности, является молекула липида, погруженная в бислой.

Функционал и обязанности
Биологическая роль мембранных белков многообразна и зависит от их структуры и расположения. Среди них выделяют рецепторные белки, канальные (ионные и порины), транспортеры, моторы и структурные белковые кластеры. Все виды мембранных белков-рецепторов в ответ на какое-либо воздействие меняют свою пространственную структуру и формируют ответ клетки. Например, рецептор инсулина регулирует поступление глюкозы внутрь клетки, а родопсин в чувствительных клетках органа зрения запускает каскад реакций, что приводят к возникновению нервного импульса. Роль мембранных белков-каналов заключается в транспорте ионов и поддержании разницы их между внутренней и внешней средой. Например, натрий-калиевые насосы обеспечивают обмен соответствующих ионов и веществ. Порины - сквозные белки - участвуют в переносе молекул воды, транспортеры - в переносе некоторых веществ против градиента концентраций. У бактерий и простейших движение жгутиков обеспечивают молекулярные белковые моторы. Структурные мембранные белки поддерживают саму мембрану и обеспечивают взаимодействие других белков плазмолеммы.
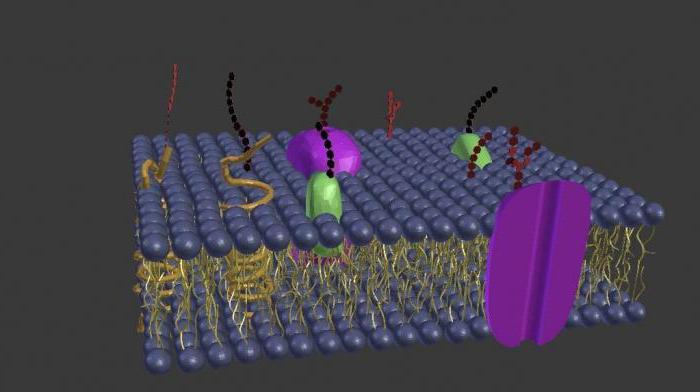
Белки для мембраны, мембрана для белков
Мембрана - это динамическая и очень активная среда, а не инертная матрица для белков, которые в ней расположены и работают. Она существенно влияет на работу мембранных белков, а липидные рафты, перемещаясь, формируют новые ассоциативные связи белковых молекул. Многие белки просто не работают без партнеров, и межмолекулярное их взаимодействие обеспечивается характером липидного слоя мембран, структурная организация которого, в свою очередь, зависит от структурных белков. Нарушения в этом тонком механизме взаимодействия и взаимозависимости приводят к нарушению функций мембранных белков и целому ряду заболеваний, таких как диабет и злокачественные опухоли.
Структурная организация
Современные представления о структуре и строении мембранных белков основаны на том, что в мембранной периферической части большинство из них состоит редко из одной, чаще из нескольких ассоциированных олигомеризующихся альфа-спиралей. Причем именно такая структура является залогом выполнения функции. Однако именно классификация белков по типам структур может принести еще немало сюрпризов. Более чем из ста описанных белков наиболее изученным по типу олигомеризации мембранным белком является гликофорин А (белок эритроцитов). Для трансмембранных белков ситуация выглядит сложнее - описан лишь один белок (фотосинтетический реакционный центр бактерий - бактериородопсин). Учитывая высокую молекулярную массу мембранных белков (10-240 тысяч дальтон), у молекулярных биологов широкое поле для исследований.

Сигнальные системы клетки
Среди всех белков плазмолеммы особое место принадлежит рецепторным белкам. Именно они регулируют, какие сигналы поступят в клетку, а какие нет. У всех многоклеточных и некоторых бактерий передача информации осуществляется посредством специальных молекул (сигнальных). Среди этих сигнальных агентов выделяют специально секретируемые клетками), небелковые образования и отдельные ионы. Последние могут выделяться при повреждении соседних клеток и запускать каскад реакций в виде болевого синдрома, главного защитного механизма организма.
Мишени для фармакологии
Именно мембранные белки являются главными мишенями применения фармакологии, так как именно они и есть те точки, через которые идет большинство сигналов. «Нацелить» лекарственный препарат, обеспечить его высокую селективность - вот главная задача при создании фармакологического средства. Избирательное воздействие только на конкретный тип или даже подтип рецептора - это влияние только на один тип клеток организма. Такое селективное воздействие может, например, отличить опухолевые клетки от нормальных.

Лекарства будущего
Свойства и особенности мембранных белков уже сегодня используются в создании лекарств нового поколения. Эти технологии основаны на создании модульных фармакологических структур из нескольких молекул или наночастиц, «сшитых» друг с другом. «Нацеливающая» часть узнает на мембране клетки определенные рецепторные белки (например, связанные с развитием онкологических заболеваний). К этой части добавляется разрушающий мембрану агент или блокатор процессов производства белков в клетке. Развивающийся апоптоз (программа собственной гибели) или другой механизм каскада внутриклеточных превращений приводит к желаемому результату воздействия фармакологического средства. В результате мы имеем лекарство с минимумом побочных эффектов. Первые такие лекарства по борьбе с раком уже проходят клинические испытания и вскоре станут залогом высокоэффективной терапии.

Структурная геномика
Современная наука о белковых молекулах все интенсивнее переходит на информационные технологии. Экстенсивный путь исследований - изучить и описать все, что можно, сохранить данные в компьютерных базах и потом искать пути применения данных знаний - такова цель современных молекулярных биологов. Всего лишь пятнадцать лет назад стартовал глобальный проект «геном человека», и мы уже имеем секвенированную карту генов человека. Второй проект, цель которого - определить пространственное строение всех «ключевых белков», - структурная геномика - пока далек от завершения. Пространственная структура определена пока только для 60 тысяч более чем из пяти миллионов белков человека. И пусть пока ученые вырастили лишь светящихся поросят и холодоустойчивые помидоры с геном лосося, технологии структурной геномики остаются этапом научного познания, практическое приложение которого не заставит долго себя ждать.


